Пол (биол.)
Пол (биол.)
Пол организмов, совокупность морфологических и физиологических особенностей организма, обеспечивающих половое размножение, сущность которого сводится в конечном итоге к оплодотворению . При этом мужские и женские половые клетки — гаметы сливаются в зиготу , из которой развивается новый организм. В зиготе объединяются 2 гаплоидных (одинарных) набора хромосом материнской и отцовской гамет. В половых клетках нового организма образуются гаплоидные наборы уже перекомбинированных (см. Рекомбинация ) отцовских и материнских хромосом (в результате обмена участками гомологичных родительских хромосом — кроссинговера — и случайного их расхождения по дочерним клеткам во время мейоза ). Поэтому в обоеполой популяции постоянно возникает множество генетически разных особей, что создаёт благоприятные условия для естественного отбора более приспособленных форм. В этом заключается основное преимущество полового размножения перед бесполым. Половое размножение преобладает у животных и высших растений; оно встречается и у многих микроорганизмов (конъюгация у бактерий сопровождается частичным обменом наследственным материалом — нитями ДНК). Половой процесс у одноклеточных организмов не требует значительной дифференциации П. (одна и та же клетка может быть и клеткой тела, и половой). У многоклеточных диплоидных организмов возникли специальные гаплоидные половые клетки: крупные и малоподвижные или неподвижные у женского П., мелкие и обычно подвижные — у мужского. У большинства растений и лишь у некоторых животных оба типа гамет производятся одной особью (см. Гермафродитизм ), у большинства животных — разными особями, которые в связи с этим строго разделяются соответственно на самок и самцов. Помимо продуцирования клеток различного П., самцы и самки различаются рядом морфологических и физиологических признаков, а также половым поведением, которые обеспечивают слияние половых клеток.
Определение пола. Все организмы, в том числе и раздельнополые, в генетическом отношении бисексуальны (двуполы), т.к. зиготы их получают генетическую информацию, потенциально дающую возможность развивать признаки мужского и женского П. У обоеполых растений и некоторых гермафродитных животных женские и мужские репродуктивные органы и половые клетки развиваются из генетически одинаковых клеток под влиянием внутренних условий (по отношению к отдельным клеткам их можно рассматривать как внешние). Механизм переключения клеток на развитие в одном случае женских, в другом мужских репродуктивных органов полностью не раскрыт. В редких случаях у раздельнополых видов потенциально бисексуальные зиготы развиваются в самок или самцов под влиянием внешних условий. Например, у морского кольчатого червя бонеллия личинка, поселяясь на хоботке самки, развивается в самца, а на дне моря — в самку. У растения Arisaema japonica из крупных клубней, богатых питательными веществами, развиваются растения с женским цветками, а из мелких клубней — с мужскими. Определение П. под влиянием внешних условий называется фенотипическим, или модификационным.
Шире распространено генетическое определение П. В этом случае зигота во время оплодотворения также получает потенциальные возможности для развития признаков обоих П. Однако под влиянием генетических факторов в одной половине зигот пересиливает тенденция развития мужского П., а в другой — женского. Специальный хромосомный механизм обеспечивает передачу одной половине потомства генов женского П., а другой — генов мужского П. В начале 20 в. было установлено, что у самцов некоторых видов насекомых в диплоидных (с двойным набором хромосом) клетках наряду с парами гомологичных хромосом имеется одна непарная хромосома. Самка же имеет две такие хромосомы. У самцов насекомых др. видов все хромосомы парные, но в одной из пар они морфологически несходные. Эти хромосомы, причастные к определению П., назвали половыми хромосомами , а остальные — аутосомами . Позднее половые хромосомы были обнаружены у многих раздельнополых организмов. Половую хромосому самца, повторяющуюся у самок, назвали Х-хромосомой, а не повторяющуюся — Y-хромосомой. Сочетание половых хромосом самца обозначают формулой X0 или XY, а самки — XX. Самцы с одной половой хромосомой продуцируют в равном количестве гаметы с Х-хромосомой и гаметы, лишённые её, т. е. с одним лишь гаплоидным набором аутосом (А); самки — гаметы только с Х-хромосомой. После случайного слияния мужских и женских гамет половина образовавшихся зигот будет иметь две Х-хромосомы (XX), а др. половина — только одну Х-хромосому. Первые станут самками, вторые — самцами (рис. ).
Самцы с разными половыми хромосомами продуцируют в равном количестве гаметы, имеющие Х-хромосому, и гаметы, имеющие Y-хромосому. Женские гаметы этого вида генетически одинаковы — все они несут по одной Х-хромосоме. В результате половина яйцеклеток будет оплодотворена сперматозоидами с Y-хромосомой, а др. половина — с Х-хромосомой. Первые зиготы, имеющие структуру XY, разовьются в особей мужского П., вторые — с XX — в особей женского П. Самцы с одной Х-хромосомой или с двумя разными (XY) хромосомами имеют гетерогаметный П., самки с ХХ-хромосомами — гомогаметный П. У многих животных, наоборот, самки имеют гетерогаметный П. Их половые хромосомы обозначают буквами Z и W или XY, а половые хромосомы гомогаметных самцов — ZZ или XX. У млекопитающих, нематод, моллюсков, иглокожих и у большинства членистоногих гетерогаметен мужской П. У насекомых и рыб гетерогаметность наблюдается как у мужского, так и у женского П. Гетерогаметность женского П. свойственна птицам, пресмыкающимся и некоторым земноводным.
Бисексуальные потенции, свойственные зиготе, обусловлены генами , локализованными в аутосомах и проявляющимися только под контролем др. генов — реализаторов П. Именно эти гены открывают путь в одном случае генам, способствующим образованию женского П., в другом — генам, обусловливающим развитие мужского П. При генетическом определении П. по типу X0, XX реализаторы женского П. локализованы в Х-хромосомах, а мужского — в аутосомах. При сочетании одной дозы реализаторов женского П., локализованных в одной Х-хромосоме, с диплоидным набором реализаторов мужского П., локализованных в аутосомах, развивается мужской П. И только 2 дозы реализаторов женского П., локализованные в 2 Х-хромосомах, пересиливают потенцию развития мужского П. и тем самым обусловливают женский П. У человека полоопределяющую роль играет Y-хромосома. В аномальных случаях она сочетается с 2, 3 и даже 4 Х-хромосомами при нормальном наборе аутосом. Хотя это и приводит к патологическим отклонениям, однако все особи с такими наборами хромосом бывают мужского П. Полоопределяющая роль Y-хромосом отмечена у многих видов животных, а среди растений — у дрёмы луговой. У дрозофилы Y-хромосома почти не содержит генов, т. е. наследственно инертна; реализаторы женского П. локализованы в Х-хромосоме, реализаторы мужского П. — в аутосомах. Развитие П. контролируется отношением Х-хромосом к набору аутосом (Х: А), условно принятым у самки за единицу (2Х:2А = 1): это отношение у самца равно 0,5 (Х:2А = 0,5). Увеличение этого отношения (полового индекса) свыше единицы приводит к чрезмерному развитию женских половых признаков («сверхсамки»), уменьшение же ниже 0,5 способствует появлению самцов с более выраженными мужскими признаками («сверхсамцы»). Особи с половым индексом 0,67 и 0,75 имеют промежуточное развитие признаков обоих П. и называют интерсексами. Явление интерсексуальности демонстрирует бисексуальную потенцию наследственной информации, передаваемой всем потомкам.
Механизм генетического контроля над развитием половых признаков может быть внутри- и межклеточным. Внутриклеточное определение П. не связано с образованием половых гормонов (например, у насекомых), и действие генов, определяющих П., ограничено клетками, в которых эти гены функционируют. При этом в одном организме могут нормально развиваться, не влияя друг на друга, участки тела с женскими и мужскими признаками (см. Гинандроморфизм ). При межклеточном определении П., характерном для млекопитающих и птиц, под контролем генов вырабатываются половые гормоны , которые, проникая во все клетки организма, обусловливают фенотипическое развитие признаков соответствующего П. Различают прогамное, сингамное и эпигамное определение П. Прогамное определение П. происходит до оплодотворения яйца, например дифференцировка яйцеклеток на быстро и медленно растущие. Первые становятся крупными, и из них после оплодотворения развиваются самки, вторые отличаются меньшими размерами и дают самцов, хотя оба вида яйцеклеток генетически одинаковы. Сингамное определение П. происходит во время оплодотворения, но на разных стадиях этого процесса. У некоторых видов с мужской гетерогаметией и физиологической полиспермией (оплодотворение яйцеклетки несколькими сперматозоидами) П. определяется в момент слияния ядер половых клеток (кариогамия ). Если с ядром яйцеклетки сливается мужское ядро с Y-хромосомой, разовьётся мужская особь, если с Х-хромосомой — женская. При женской гетерогаметии П. потомства зависит от того, какая из половых хромосом попадает в ядро яйцеклетки во время мейоза. Если в ядре окажется Z-хромосома, разовьётся особь мужского П., если W-хромосома — женского. Т. о., в данном случае П. зиготы устанавливается до кариогамии. Эпигамное определение П. наблюдается у разнополых видов с фенотипическим определением П., когда направленность развития в сторону мужского или женского П. обусловливается влиянием внешних условий после оплодотворения.
Зависимость признаков от пола . Зависят от П. признаки, ограниченные и контролируемые им. Ограниченные П. признаки в силу половой дифференциации могут проявиться только у одного из П. (продукция молока или яиц свойственна только женскому П.), хотя полимерные гены этих признаков локализованы в аутосомах обоих П. Признаки, контролируемые П., проявляются или у обоих П. (с разной степенью выраженности), или (чаще) только у одного из П. (более мощное развитие рогов у баранов, бороды — у козлов), хотя оба в равной мере содержат в аутосомах гены этих признаков. Несходное их развитие обусловлено значительным различием физиологических процессов в организмах разного П.
Гены, детерминирующие признаки, сцепленные с П., локализованы как в парных, так и непарных половых хромосомах и поэтому наследуются иначе, чем признаки, обусловленные парными генами, локализованными в аутосомах обоих П. Если гены локализованы в непарной Y-хромосоме гетерогаметного самца, то обусловливаемые ими признаки наследуются лишь сыновьями, а при локализации генов в хромосоме гетерогаметной самки — только дочерьми. Наследуемые т. о. признаки называются голандрическими. Этот тип наследования обнаружен у некоторых видов рыб и насекомых. У др. видов животных он с полной достоверностью не доказан. При локализации генов в гомологичных Х- или Z- хромосомах обусловленные ими признаки передаются сцепленно с П. по типу, получившему название наследования крест-накрест, когда рецессивный признак матери проявится у сыновей, а доминантный — у дочерей (Т. X. Морган), что встречается у многих видов животных (например, трёхцветность кошек, полосатость окраски оперения и скорость его роста у кур). Много сцепленных с П. мутаций обнаружено у дрозофилы и тутового шелкопряда.
Сцепленными с П. могут быть и летали — гены, обусловливающие смертельный исход при развитии организма. Если гомогаметный родитель гетерозиготен по летали, локализованной в одной из гомологичных половых хромосом (X или Z), то половина его гетерогаметных потомков погибнет, получив деталь, губительному действию которой в генотипе не будет противопоставлен нормальный аллель . При гетерогаметии женского П. от леталей гибнет половина дочерей, а при гетерогаметии мужского П. — половина сыновей. Иногда мутантные гены в Х- и Z- хромосомах лишь частично снижают жизнеспособность потомства или вызывают различные заболевания, наиболее часто проявляющиеся у гетерогаметного П. У человека обнаружено свыше 50 сцепленных с П. мутаций, приводящих большей частью к нарушению нормальной жизнедеятельности организма (см. Хромосомные болезни ).
Соотношение полов . При фенотипическом определении П. оно зависит от количества развивающихся организмов, которые попадают под влияние внешних факторов, детерминирующих тот или иной П. При генетическом определении П. соотношение П. у большинства видов, как правило, очень близко к 100?: 100? (100 самок: 100 самцов). Однако и при таком определении П. есть отклонения. Так, у некоторых видов млекопитающих с мужской гетерогаметией статистически достоверно рождается на 1—2% больше потомков мужского П.
Регуляция пола . Существенный сдвиг соотношения организмов в сторону одного из П. имеет как теоретическое, так и практическое значение, т.к. один из П. обычно более продуктивен. Методы регуляции П., сведённые к 4 основным направлениям, применяются в зависимости от типа определения П. и биологических и хозяйственных особенностей вида.
Фенотипическое переопределение П. Если действие генов П. реализуется посредством гормонов, половые признаки изменяются при пересадке половых органов одного П. другому (см. Гонады ) или при введении в организм гормонов противоположного П., а также некоторых аминокислот. Степень фенотипических изменений П. зависит от особенностей вида и дозы введённого препарата. Однако лишь в редких случаях (у некоторых рыб и земноводных) особи с фенотипически переопределённым П. продуцируют гаметы, противоположные их генотипическому П. В следующем поколении, если действие гормонов прекращается, снова вступает в силу генетический механизм определения П.
Управление генетическим механизмом определения П., или искусственное сочетание в яйцеклетке половых хромосом. Направленное изменение соотношения П. достигнуто в экспериментах с тутовым шелкопрядом, у которого П. строго определяется сочетанием половых хромосом (ZW — ?; ZZ — ?). Неоплодотворённые яйца после прогрева развиваются партеногенетически (см. Партеногенез ) за счёт диплоидного ядра, не завершившего редукционного деления. Все клетки партеногенетического эмбриона сохраняют материнскую структуру, в частности и в отношении половых хромосом ZW, и, следовательно, развиваются только в самок (Б. Л. Астауров). Воздействием ионизирующих излучений и прогревом удалось подавить в свежеотложенном осеменённом яйце женское ядро и переключить развитие на мужское начало. Диплоидное ядро мужской зиготы образуется путём слияния двух мужских ядер и поэтому имеет структуру мужского П. ZZ. Из таких зигот развиваются гусеницы всегда мужского пола (X. Хасимото; Б. Л. Астауров). Этими методами впервые у с.-х. вида шелкопряда решена проблема произвольной регуляции П. У млекопитающих учёные пытаются разделить по морфологическим и физиологическим особенностям Х- и Y-сперматозоиды с целью последующего осеменения одной категорией сперматозоидов. Однако этим способом пока не удалось достоверно сместить соотношение П.
Раннее распознавание П. Используется для сортировки вылупившихся цыплят на петушков и курочек по окраске оперения, сцепленной с П., а также для «сверхранней» сортировки по П. тутового шелкопряда. Под действием ионизирующего облучения у шелкопряда пересажена аутосома с доминантным геном, обусловливающим тёмную окраску яиц тутового шелкопряда, на половую W- хромосому. Сцепление хромосом стойко передаётся по наследству. Те яйца, в которые попадает W- хромосома с пересаженным доминантным геном, приобретают тёмный цвет и развиваются в самок, в то время как яйца мужского П., не получив доминантного гена, остаются непигментированными. Фотоэлектрические автоматы с большой скоростью разделяют разноокрашенные яйца по П. Выведенные таким способом (В. А. Струнников и Л. М. Гуламова) меченые по П. породы шелкопряда находят практическое применение в советском шелководстве. В 60-х гг. 20 в. в опытах английских учёных Р. Эдуардса и Р. Гарднера зафиксировано рождение потомства только одного П. и у млекопитающих. У кроликов извлекали из тела матери ранних зародышей, цитологическим методом определяли их П. и затем зародышей нежелательного П. выбраковывали, а зародышей нужного П. возвращали в матку. Около 20% возвращенных зародышей прижилось и развивалось в крольчат предсказанного учёными П.
Изменение соотношения П. может быть почти у всех животных с генетическим определением П. результатом гибели половины зародышей гетерогаметного П. под действием сцепленных с П. деталей. Однако для многих с.-х. животных такой подход к регуляции П. экономически не оправдан. Исключение составляет тутовый шелкопряд. В СССР радиационным методом выведена (В. А. Струнников) генетически особая порода тутового шелкопряда, у которой в обоих Z- хромосомах самцов всегда имеется по одной негомологичной друг другу летали (сбалансированные летали). Если этих самцов скрестить с самками обычных пород, на стадии яйца одна половина самок погибнет от первой, а другая — от второй летали. Из яиц мужского П. вылупляются нормальные гусеницы. Этот способ позволяет в неограниченных количествах получать у тутового шелкопряда только один более продуктивный мужской П.
Эволюция пола . Раздельнополость (бисексуальность), свойственная уже многим одноклеточным (водоросли, простейшие), произошла от смешаннополости. Лишь в некоторых случаях (например, при паразитизме ) смешаннополость могла возникнуть вторично из раздельнополости. Так, у паразитических ракообразных наблюдаются все переходы от смешанно- к раздельнополости (например, раздельнополые виды с хорошо развитыми самками и карликовыми самцами — явственный сдвиг в сторону гермафродитизма). Фенотипическое определение П. древнее генетического, т.к. на ранних стадиях эволюции ещё не существовало специального аппарата половых хромосом. Возникающие на определённых этапах эволюции специальные половые хромосомы (рыбы, земноводные) первоначально морфологически неотличимы от аутосом, и о наличии их можно судить только по признакам, сцепленным с П. Вслед за морфологическими различиями между половыми хромосомами и аутосомами возникает дифференцировка между Х- и Y- хромосомой, что делает всё более редкой конъюгацию между ними и затрудняет обмен их участками при кроссинговере (перекресте). Всё это способствует выполнению специфической функции половых хромосом — быть реализаторами женского или мужского П. Полное исчезновение Y- хромосом делает генетическое определение П. ещё более совершенным: П. определяется равновесием между числом аутосом и хромосом.
Лит.: Астауров Б. Л., Генетика пола, в сборнике: Актуальные вопросы современной генетики, [М.], 1966; Бреславец Л. П., Определение и наследственность пола у высших растений, «Тр. по прикладной ботанике, генетике и селекции, Сер. 2», 1934, № 6; Рыжков В. Л., Генетика пола, [Хар.], 1936; Лобашев М. Е., Генетика, 2 изд., Л., 1967.
В. А. Струнников.
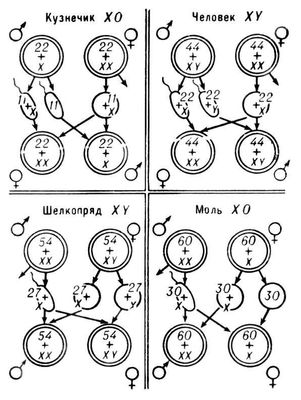
Схема различных типов хромосомного определения пола (двойные кружки — зиготы; одинарные — гаметы; цифры указывают число хромосом).
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Пол (биол.)
Пол (биол.) Пол организмов, совокупность морфологических и физиологических особенностей организма, обеспечивающих половое размножение, сущность которого сводится в конечном итоге к оплодотворению . При этом мужские и женские половые клетки — гаметы сливаются в
Плод (биол.)
Плод (биол.) Плод (fetus), млекопитающее животное или человек в утробный период развития после закладки основных органов и систем: у человека — это период начиная с 9-й недели и до момента рождения. На 9-й неделе развития П. по внешнему виду принимает черты тела человека: ясно
Класс (биол.)
Класс (биол.) Класс (от лат. classis — разряд, группа) (биологическое), одна из высших таксономических (систематических) категорий животных и растений. К. объединяет родственные отряды (животных) или порядки (растений). Например, отряды грызунов, насекомоядных, хищных и др.
Сон (биол.)
Сон (биол.) Сон, периодическое физиологическое состояние мозга и организма человека и высших животных, внешне характеризующееся значительной обездвиженностью и отключением от раздражителей внешнего мира. Субъективно у человека при этом угнетается осознаваемая